Notes � Andy King, hearing
Greg Detre
Tuesday, 04 December, 2001
Neuro II, essay VI
Dr Andrew King
Essay title
How are auditory
stimuli encoded at different levels of the central auditory pathway?
Notes � Kandel & Schwarz, Principles 3rd edn, ch 32 on �Hearing�
Ohm suggested 100 years ago that the ear and brain deconstruct use Fourier analysis to deconstruct/simplify complex waveforms into the sum of many (simpler) individual sine and cosine waves of appropriate frequencies, phases and amplitudes.
Sound is produced by vibrations that result in the alternating compression and rarefaction (increased or decreased pressure) of the surrounding air.
Speed of sound in air at standard temperature/pressure = ???
Sound is produced by vibrations and is transmitted through air by pressure waves
Vibrations of hair cells are transformed into electrical signals in the auditory nerve
Summary
�The external ear and middle ear form collectively a mechanical transmission system that converts sounds, or air pressure waves, into fluid waves in the inner ear.�
�The receptors of the inner ear, the hair cells, act like miniature amplifiers, each tuned mechanically to provide a maximal electrical response when vibrated at a particular frequency by the fluid waves of the inner ear. The hair cells are a set of frequency filters ordered spatially within the cochlea; those with high-pass frequencies occupy the bass, and those with low-pass frequencies occupy the apex. Sensory transduction occurs in the organ of Corti, where the hair cells interact with supporting elements to convert fluid waves into the bending of the hair bundles and resultant ion influxes. The organ itself is under dynamic control from the brain, so it may be tuned to sounds of particular interest.�
�Signal coding occurs initially at the synapse between the hair cells and the fibres of the auditory nerve. Using only the digital code of action potentials, the nerve provides a profile of sound input, including the spectrum of frequencies, the phase or timing relations of different frequency components, and their relative amplitudes. Given the complexity of the information encoded, it is not suprising that this code is not understood completely. It seems clear, howerver that there is a relationship between the site a nerve fibre innervates in the cochlear and the frequency characteristics of the fibre. Thus, each fibre responds best to a very narrow band of frequencies, although most fibres are excited to some extent by a wide range of frequencies.�
�In the brain, inputs from the two ears are combined by ascending pathways that cross the midline extensively. The pathways separate information about the timing and intensity of signals, the binaural cues for sounds localisation. The information ascends in parallel to the auditory cortex where the timing, intensity and frequency of sound are mapped. The diversity of separate areas within the auditory cortex reflects the complexity of the task underlying perception of complex sounds. As in the visual cortex, � the auditory cortex[�s] separate functional regions deconstruct speech into components to generate a perception of location, loudness and pitch.�
Notes � Pickles, �An introduction to the physiology of hearing�
Ch 4, �The auditory nerve�
Summary, pg 109
Most of the fibres in the auditory nerve innervate hair cells
Fibres� tuning curves (the relation between threshold and stimulus frequency) vary, with some having lower thresholds to tones of some frequencies than of others, and each having a �characteristic frequency� (its threshold minimum, ).
Fibres show a sigmoidal relation between firing rate and stimulus intensity.
The frequency resolving power (how selective for a given frequency it is) has been measured by a �quality� factor, calculated as the characteristic frequency divided by the bandwidth of the fibre to tones at an intensity 10 dB abovce the best threshold (known as �Q10�). Fibres with a high Q10 have good frequency selectivity, e.g. the cat�s greatest Q10s are about 8 at about 10kHz.
�During tonal stimulation, auditory nerve fibres fire preferentially during one part of the cycle of the stimulating waveform if the stimulus is below 4-5kHz. The fibres are excited by deflection of the basilar membrane in only one direction.�
�For fibres with characteristic frequencies below 4-5kHz, clicks preferentially evoke responses at certain intervals after the stimulus��
�One tone can reduce, or suppress, the response to another [�two-tone suppression�], even though single tones are only excitatory�. This arises from the non-linear properties of the basilar membrane mechanics, and works for stimuli other than tones too. Also, a stimulus can mask another if it produce a greater firing rate.
Ch 7 on �The auditory cortex�
Basic anatomy
Summary, pg 232
The auditory cortex consists of a:
�core� area � the primary auditory cortex, AI, which receives it input from the main specific auditory relay of the thalamus, the ventral division of the medial geniculate body)
surrounded by a �belt� � receives its input mainly from the other divisions of the medial geniculate
The primary auditory cortex, and some divisions of the belt area, are tonotopically organised. Iso-frequency strips lie at right angles to the line of frequency progression. However, there do not appear to be sudden jumps in frequency as an electrode is moved tangentially in the cortex.
Binaural dominance does seem to be related to the existence of discrete columns (with cells of the same binaural dominance lying in the same radial direction in the cortex, and segregated into discrete strips, running along the cortical surface at roughly right angles to the iso-frequency strips).
Not all neurons in the primary auditory cortex show responses to sound.
There are various shapes of tuning curves in the primary auditory cortex (e.g. broad, multi-peaked), with complex temporal patterns of response.
Many show binaural interactions, suggesting that they code for sound direction. Each cortex predominantly represents sound sources on the contralateral side.
Some cells seem specifically responsive to frequency-modulated stimuli. Othres respond only to complex sounds such as animal calls (though they are not specific detectors for these so much as responding to their basic acoustic elements).
Cortical lesion studies
Initially seemed to show that frequency discrimination is impossible after complete lesions of the auditory cortex. However, frequency discrimination in a series of ongoing pips appears to be possible, while other forms of frequency discrimination are not, apparently purely as a result of task difficulty.
AI seems heavily implicated in sound localisation. Sound locus is coded in a frequency-specific way, such that each iso-frequency strip is involved in coding the source locus for sounds of that frequency.
Upset tasks where the animals have to utilise the temporal dimension of auditory stimuli, or detect/discriminate very short stimuli. The auditory cortex may be necessary for auditory short-term memory, and prolonging the effects of short stimuli.]
The auditory cortex seems to affect he ability to attend to sounds in the contralateral ear.
Hypotheses about auditory function:
necessary for the analysis of complex sounds
subserves sound localisation and the representation of �auditory space�
necessary for selective attention to auditory stimuli on the basis of source position
serves to inhibit inappropriate motor responses
serves to identify stimuli on an absolute basis
necessary for the discrimination of auditory temporal patterns
necessary for short-term memory when one auditory stimulus has to be related to another later in time
necessary for difficult auditory tasks
Ch 8 on �The centrifugal pathways�
Basic anatomy
�The centrifugal auditory pathways run from the higher stages of the auditory system to the lower. One pathway, the olivocochlear bundle, runs from the superior olivary complex to the hair cells of the cochlea. The central auditory nuclei are targets for other centrifugal pathways. It has been suggested that the pathways are organised into a chain, running from the cortex to the cochlea.�
Summary
�Centrifugal (efferent) auditory pathways parallel the centripetal (afferent) auditory pathways along the entire length of the system, forming a chain which runs from the cortex to the hair cells�, often running �adjacent to, but not actually within, the tracts and nuclei principally associated with the ascending system�.
�The cochlea is innervated by the olivocochlear bundle, which arises bilaterally in the superior olivary complex. Cells of a medial group, situated near the medial nucleus of the superior olive, have relatively large bodies, and give rise to relatively large axons which innervate the outer hair clels, mainly on the contralateral side. Cells of a lateral group are situated around the lateral nucleus of the superior olive, have smaller cell bodies, and give rise to smaller axons which innervate the nerve fibres just below the inner hair cells, almost entirely on the ipsilateral side.�
Electrical activation of the crossed olivocochlear bundle reduces the response of auditory nreve fibres to sound, perhaps because the efferents affect the outer hair cells� signals by affecting the tuning and sensitivity of the mechanical travelling wave on the basilar membrane.
Fibres of the olivocochlear bundle are themselves responsive to sound, and have sharp tuning curves and low thresholds like the afferent fibres. They terminate in corresponding areas of the cochlea , making closed frequency-specific feedback loops. Their activity is also affected by central influences.
Possible functions of the olivocochlear bundle in auditory performance:
�may improve the detection of signals in masking noise
may help protect the cochlear from acoustic trauma
may control the mechanical state of the cochlear, compensating for changes in factors like stiffness + static displacemtn foth ebasilar membrance
may be involved in attention�
�The cochlear nucleus receives branches of the olivocochlear bundle, toghetherw other centrifugal fibres from the superior olivary complex, and from higher auditory nuclei, including the nuclei of the lateral lemniscus and the inferior colliculus.�
Centrifugal fibres to the cochlear nucleus are both inhibitory and excitatory. It affects the ability to detect signals in noise, and the bandwidth of noise that contributes to masking.
Pickles, �An introduction to the physiology of hearing�
Notes � Instant notes neuroscience
Peripheral auditory processing
Primary auditory afferents � Cell bodies in the spiral ganglion and send their central axons to the pons via the eighth cranial nerve. They fire spontaneously, and increase their firing in response to a tone. Most are sharply tuned.
Frequency coding � two ways:
place coding � for frequencies above 3000Hz, the frequency response of an afferent depends on where along the basilar membrane it is from (tonotopic = frequency-position mapping)
temporal coding � for lower frequencies, afferents fire during a particular phase of the waveform (phase-locking). within the population, each afferent only needs to fire occasionally.
Sound level � auditory afferents respond only to a limited range of sound pressure levels (SPLs). The full range is encoded by populations of afferents with different dynamic ranges. The most sensitive afferents have the highest spontaneous firing rates. Efferents from the superior olivary complex innervate hair cells, reducing their sensitivity, enabling them to respond to high sound levels.
Central auditory processing
Central auditory pathways � Primary auditory afferents terminate in the cochlear nuclei in the pons. Ventral cochlear axons go to the superior olivary complex on both sides. This projects to the nuclei of the lateral lemniscus, and mainly deals with sound localisation. The dorsal cochlear nucleus projects directly to the contralateral nucleus of the lateral lemniscus. The nuclei of the lateral lemniscus sends axons to the inferior colliculus of the tectum, which projects to the medial geniculate nucleus (MGN). This goes to AI, and is responsible for conscious sound perception. Though the largest auditory pathway is contralateral, extensive connections across the midline ensure interactions between sides.
Cochlear nuclei � Different cell types process different features. Bushy cells signal exact timing information to the (binaural) medial superior olivary nucleus for sound localisation. Stellate cells signal sound level. Many cells are frequency-tuned, further tuned by lateral inhibition.
Tonotopic mapping � Systematic frequency representation (all frequencies are roughly equally represented in humans). In AI, there are isofrequency columns perpendicular to cortical surface, arranged in tonotopic bands.
Notes � Kaas, Hackett and Tramo (1999), �Auditory processing in primate cerebral cortex� in Current opinion in Neurobiology
Abstract
�Auditory information is relayed from the ventral nucleus of the medial geniculate complex to a core of three primary or primary-like areas of auditory cortex that are cochleotopically organised and highly responsive to pure tones. Auditory information is then distributed from the core areas to a surrounding belt of about seven areas that are less precisely cochleotopic and genearlly more responsive to complex stimuli than tones. Recent studies indicate that the belt areas relay to the rostral and caudal divisions of a parabelt region at a third level of processing in the cortex lateral to the belt. The parabelt and belt regions have additional inputs from dorsal and magnocellular divisions of the medial geniculate complex and other parts of the thalamus. The belt and parabelt regions appear to be concerned with integrative and associative functions involved in pattern perception and object recognition. The parabelt fields connect with regions of temporal, parietal and frontal cortex that mediate additional auditory functions, including space perception and auditory memory.
Notes � web
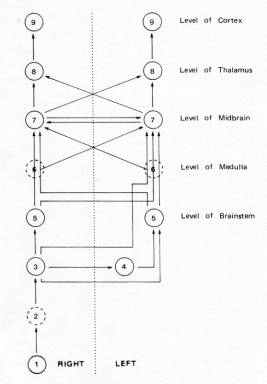

9: Primary
Auditory Cortex
8:� Medial Geniculate Body
7:� Inferior Colliculus
6:� Lateral Lemniscus
5:� Superior Olivary Complex
4:� Trapezoid Body
3:� Cochlear Nucleus
2:� Auditory Nerve Fibers
1:� Cochlear Hair Cells
Excerpt from sound localisation essay
Romanski et al examined the connectivity of higher auditory cortical areas in macaque monkeys, using a combination of anatomical tracer dyes with electrophysiological recordings. Their results support the ventral/dorsal temporal/parietal what/where processing dichotomy, contributing to functionally distinct regions of the frontal lobe. Further parallelism is evident in the primate auditory cortex, which has three similar primary or primary-like areas, each tonotopically organised and receiving activating inputs directly from the auditory thalamus. The projections from these to 7 or 8 proposed fields seem to provide anatomical support for the beginnings of ventral and dorsal cortical processing streams - they project to largely different portions of the frontal lobe. However, the middle belt area makes connections to the frontal lobe that overlap those of the two putative streams, indicating possible intermediate or additional auditory streams (also analogous to the additional functional streams or 'streams within streams' found in the visual system).
Questions
what�s the difference between sound and all the random motion of the air??? if the answer is �nothing at all�, why don�t we hear a constant cacophony�???
surely because even though the energy of a sound wave is very small, the overall motion of the molecules in the air, like white noise, is not concerted enough to be noticeable
what�s white/pink noise, spectrum???
�random: waveform varies randomly in amplitude over time; amplitude normally distributed around the mean.
white noise: spectrum flat over frequency
pink noise: spectrum energy declines as a function of frequency�
which bits are tonotopic, which aren�t??? how do the tonotopic representations vary???
to what extent does cell type relate to cell function???
which lobe is AI in???
topographic vs anatomic??? correspond to representation and physiology???
how do the various tonotopic representations at the different levels???
what about the descending connections from cortex to thalamus???
Kandel & Schwarz
Pickles
�Single fibres of the auditory nerve are always excited by auditory stimuli and never show sustained inhibition to single stimuli� � eh??? (pg 110)
core/belt = primary/secondary
Instant notes
what/where is the spiral ganglion???
Kaas, Hackett and Tramo
no difference between cochleotopic and tonotopic, is there???
is AII tonotopically organised???